Type I interferons (IFNs) drive antiviral immunity through ISG induction, yet transcript levels often do not reflect actual protein activity. This article outlines why transcript–protein gaps occur and how direct IFN measurement improves insight.
Type I interferons (IFNs), including IFN‑α subtypes and IFN‑β, orchestrate antiviral immunity through JAK‑STAT signaling and ISG induction, which has made transcriptomic profiling central for mapping canonical and non‑canonical IFN pathways [1,2] and for defining disease endotypes in IFN‑I‑driven pathogenesis [3,4]. However, transcript levels frequently diverge from true protein abundance or function due to post‑transcriptional regulation, translational efficiency, and degradation dynamics, and studies show that the IFN gene signature (IGS) cannot fully substitute for direct cytokine measurements [5,6]. This article examines the biological basis of transcript–protein discrepancies and highlights how high‑sensitivity ELISA and direct IFN protein quantification complement transcriptomics to deliver more complete mechanistic insight.
Understanding the Transcript-Protein Disconnect in Type I IFN Biology
Viral infection triggers cells to synthesize Type I interferons—predominantly IFN-α and IFN-β—which subsequently engage the interferon-α receptor (IFNAR) displayed on cell surfaces [3]. IFNAR engagement initiates JAK-STAT signaling cascades, activating transcription factors that induce hundreds of ISGs [1,2]. The resulting ISG products construct a multifaceted antiviral barrier: they degrade viral RNA, obstruct viral protein synthesis, inhibit viral genome replication, and fine-tune immune responses [2].
(IFNAR) displayed on cell surfaces [3]. IFNAR engagement initiates JAK-STAT signaling cascades, activating transcription factors that induce hundreds of ISGs [1,2]. The resulting ISG products construct a multifaceted antiviral barrier: they degrade viral RNA, obstruct viral protein synthesis, inhibit viral genome replication, and fine-tune immune responses [2].
Genome-wide analyses have established that mRNA abundance accounts for about 40% of the variation observed in protein levels [7,8]. This transcript-protein discrepancy becomes particularly acute in Type I interferon biology. Detailed investigations of immune response dynamics have documented substantial mismatches between transcript induction patterns and actual protein accumulation [9,10]. The implication is important: ISG transcript measurements alone obscure the functional reality, as numerous transcriptionally activated ISGs fail to generate proportional protein responses.
What drives this biological disconnect? Several mechanisms operate simultaneously:
- Differential Translation Efficiency: ISG mRNAs exhibit markedly unequal translation rates [10,11]. Numerous ISG transcripts harbor regulatory elements that modulate how efficiently they're translated into protein. Consider IRF7, the interferon regulatory factor essential for amplifying IFN production—it experiences selective translational repression by other ISG products, creating an unexpected regulatory loop.
- Disparate Protein Stability: While ISG mRNAs typically persist for several hours, their protein products demonstrate remarkable stability differences spanning minutes to days [9]. Numerous interferon signaling proteins undergo rapid, precisely controlled degradation via ubiquitin-proteasome pathways. This creates temporal control over signaling independent of ongoing mRNA transcription.
- Post-Translational Regulation: Phosphorylation, ubiquitination, and related modifications dramatically alter protein function and longevity [11]. STAT1 exemplifies this principle—phosphorylation converts it from a transcriptional repressor to an activator [3]. This functional switch remains completely invisible to transcript-based measurements.
- Feedback Inhibition of Translation: In a paradoxical twist, certain ISG products actively suppress translation as part of their antiviral mechanism [2]. This creates regulatory circuits where initial ISG expression dampens translation of subsequently induced ISG transcripts.
The Imperative for Immunoassay-Based Type I IFN Quantification
Quantifying Functional Molecules Directly
Immunoassays detect actual protein molecules present in biological specimens rather than cellular productive capacity. For Type I interferons, this distinction carries profound significance. Circulating IFN-α and IFN-β exist at remarkably low concentrations—typically picograms per milliliter in serum or plasma [12,13]. These concentrations represent the bioactive molecules that physically engage IFNAR receptors and trigger downstream signaling cascades. Immune cell transcript measurements cannot predict these circulating protein concentrations for several reasons:
- IFNs are secreted proteins whose extracellular concentrations depend on production rates, secretion efficiency, and systemic clearance
- Distinct cell types secrete IFNs at different rates despite comparable transcript levels
- Autocrine and paracrine signaling generates local IFN concentrations that diverge substantially from systemic measurements
- Both viral and host factors can obstruct IFN secretion while leaving transcription intact
Sensitivity for Scarce Proteins
Contemporary high-sensitivity ELISA platforms detect Type I interferons at concentrations in the low picogram-per-milliliter range [12,13]. This translates to measuring mere hundreds of molecules within small sample volumes. Such sensitivity proves essential because circulating IFN levels remain quite low even during active viral infections. Healthy individuals typically show undetectable IFN or concentrations in the low single-digit pg/mL range.
Modest IFN increases (mere 10-20 pg/mL changes) can fundamentally shift cellular states from quiescent to robustly activated [3]. Capturing these biologically consequential changes demands the sensitivity that immunoassays uniquely deliver. Type I IFN production also exhibits rapid kinetics during infection: concentrations peak within hours of viral exposure before declining over 24-48 hours. Serial immunoassay measurements can track these dynamic temporal patterns.
Accurate Measurement of IFN Species
The Type I interferon family encompasses multiple IFN-α subtypes—at least 12 in humans—plus IFN-β, each possessing distinct biological activities [2]. Even when subtypes appear to induce similar ISG signatures at high doses, their differences in receptor binding and natural physiological concentrations still lead to meaningful functional diversity. A High-quality immunoassay that can accurately measure total amount of IFN‑α subtypes becomes essential. 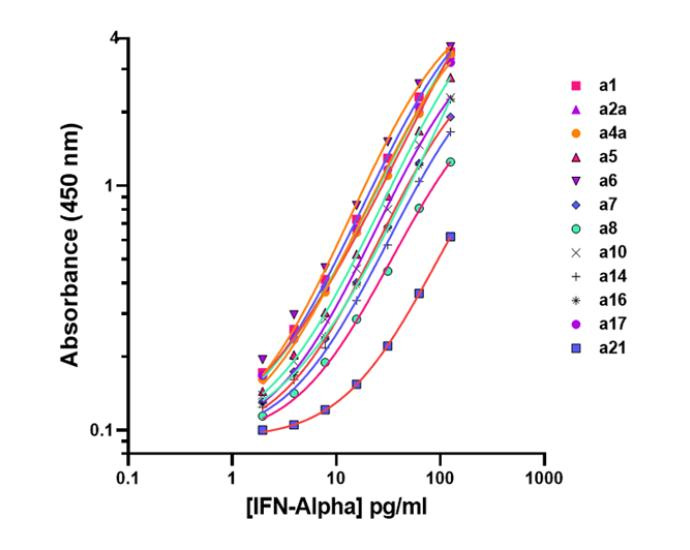
A key reason total IFN‑α measurement is essential is that human diseases (including autoimmunity, viral infections, and cancer) produce complex mixtures of IFN‑α subtypes, each contributing differently to inflammation, antiviral defense, and overall immune tone. These conditions often generate distinct and shifting subtype expression patterns, making any single‑subtype readout an incomplete and potentially misleading indicator of IFN‑α–driven biology. Because all IFN‑α subtypes ultimately signal through the same IFNAR pathway, what matters biologically is the aggregate interferon activity produced by all 12 subtypes together. For this reason, the combined concentration of total IFN‑α provides the most accurate reflection of systemic type I interferon signaling, offering a more reliable and clinically meaningful biomarker than measuring any individual subtype alone.
Clinical Sample Compatibility
High-sensitivity ELISA quantifies Type I IFNs using just 100 μL of serum or plasma, making it compatible with standard clinical blood collection. In vitro studies measure secreted IFN in culture media, directly assessing cellular IFN production capacity. Immunoassays also quantify IFNs in cerebrospinal fluid, synovial fluid, bronchoalveolar lavage fluid, and tissue homogenates, revealing local IFN production at infection or inflammation sites.
Clinical Applications where Immunoassay-Based IFN Quantification Matters
Autoimmune Disease Applications
- Systemic Lupus Erythematosus (SLE): The interferon signature in SLE—characterized by elevated ISG transcript expression in peripheral blood—correlates with disease activity across many patients [3,4]. However, direct circulating IFN-α and IFN-β measurement via immunoassay reveals heterogeneity: some patients with elevated ISG transcripts show normal or modestly elevated IFN protein levels, while others demonstrate marked protein elevation [5,6]. This distinction carries clinical weight: patients with elevated circulating IFN proteins appear to respond more favorably to anti-IFN therapies such as anifrolumab [14]. Serial IFN protein measurements track disease activity and treatment responses more directly than transcript-based signatures.
- Type I Interferonopathies: These rare genetic disorders—including Aicardi-Goutières syndrome and STING-associated vasculopathy—result from mutations causing constitutive IFN production. Patients demonstrate sustained circulating IFN-α elevation measurable by high-sensitivity immunoassay [12]. IFN protein concentrations correlate with disease severity and neurological manifestations, rendering immunoassays essential for both diagnosis and monitoring.
Viral Infection Contexts
- Acute Viral Infections: COVID-19 patient studies have demonstrated that absent or delayed IFN responses associate with severe disease progression. Critically, some patients with severe illness exhibited low or undetectable circulating IFN despite robust ISG transcript induction—a disconnect that transcriptomics miss entirely but immunoassays readily detect. Immunoassay-based circulating IFN-α and IFN-β measurement reveals response timing and magnitude, individual production capacity variation, and viral IFN suppression strategies.
- Chronic Viral Infections: Chronic hepatitis C, HIV, and hepatitis B infections demonstrate complex interferon system dysregulation. Immunoassay-based monitoring establishes baseline IFN status before antiviral therapy, predicts treatment responses, and assesses IFN-related toxicity during therapy.
Cancer Immunotherapy Implications
Tumor microenvironment Type I IFNs can enhance anti-tumor immunity by activating dendritic cells and cytotoxic T lymphocytes [2]. Immunoassay-based IFN measurement in tumor interstitial fluid or lysates assesses local IFN activity and correlates with checkpoint inhibitor responses. For therapeutic IFN-α applications in specific malignancies, immunoassay-based monitoring verifies therapeutic IFN delivery, enables pharmacokinetic profiling, and detects anti-drug antibodies.
Type I IFN Pathway Therapeutics
Monoclonal antibody blocking the IFNAR receptor has gained approval for SLE treatment [14]. Maximal efficacy occurs in patients with elevated baseline IFN activity, and immunoassay-based circulating IFN measurement helps identify patients most likely to benefit. For emerging therapeutics that directly neutralize IFN-α or IFN-β proteins, immunoassay-based pharmacodynamic monitoring quantifies free versus drug-bound IFN and detects emerging anti-drug antibodies.
Immunoassays and Transcriptomics: A Complementary Framework
Immunoassay-based IFN quantification and transcript-based methodologies provide complementary information. Transcriptomics excels at comprehensive ISG profiling—simultaneously measuring hundreds of genes [2], revealing cellular transcriptional states, and enabling novel IFN-regulated gene discovery. Immunoassays excel at precise functional IFN protein quantification, detecting low-abundance proteins at physiological concentrations in clinically accessible samples.
Optimal approaches integrate both modalities [11]. In SLE research, ISG transcriptional signatures identify patients with activated IFN pathways, while immunoassay-based measurement determines which patients show elevated IFN proteins [5,6]. Some patients demonstrate ISG transcript elevation without elevated circulating IFN (suggesting paracrine or intracellular IFN activity), while others show elevated circulating IFN despite modest ISG signatures. This integrated perspective enables patient stratification impossible with either modality alone.
Conclusions
The transcript-protein abundance gap presents a fundamental challenge in Type I interferon research, carrying significant implications for disease mechanism understanding and biomarker development [7,8,11]. While transcriptomic approaches offer unmatched breadth for cellular response profiling, they cannot substitute for direct measurement of functional IFN proteins driving biological outcomes.
Immunoassay-based quantification, particularly high-sensitivity ELISA, represents the measurement gold standard for Type I interferons. Modern immunoassay exceptional sensitivity (picogram-per-milliliter detection) [12,13], clinical sample compatibility, and IFN subtype discrimination make them indispensable for research and clinical applications.
For researchers, immunoassays provide IFN protein level ground truth that complements and validates transcriptomic findings. For translational applications, IFN immunoassays enable patient stratification, treatment selection, and therapeutic monitoring across autoimmune diseases, viral infections, and cancer immunotherapy [14-16]. Understanding Type I interferon responses demands measuring what actually exists rather than what cells are programmed to produce. Direct IFN protein measurement enables complete interferon response understanding, clinical outcome prediction, and effective IFN-targeted therapy development.
References:
- Mazewski, C. et al., (2020), "Type I Inteferon (IFN)-Regulated Activation of Canonical and Non-Canconical Signaling Pathways", Front Immunol., 11:606456, DOI: 10.3389/fimmun.2020.606456 (link)
- Schneider WM., et al., (2014), "Interferon-stimulated genes: a complex web of host defenses", Annu Rev Immunol., 32:513, DOI: 10.1146/annurev-immunol-032713-120231 (link)
- Ivashkiv, LB. et al., (2014), "Regulation of type I interferon responses", Nat Rev Immunol., 14(1):36, DOI: 10.1038/nri3581 (link)
- Cataloma, MD. et al., (2019), "Gene expression analysis delinates the potential roles of multiple interferons in systemic lupus erythematosus", Commun Biol., 2:140, DOI: 10.1038/s42003-019-0382-x (link)
- Winthrop, K. et al., (2024), "The intereferon gene signature as a clinically relevant biomarker in autoimmune rehumatic disease", Lancet Rheumatol., 6(3):e195, DOI: 10.1016/s2665-9913(21)00254-x (link)
- Wright HL. et al., (2022), "Interferon-a-mediated therapeutic resistance in early rheumatoid arthritis implicates epigenetic reprogramming", Ann Rheum Dis., 81(9):1414, DOI: 10.1136/annrhemdis-2022-222370 (link)
- Schwanhausser B. et al., (2011), "Global quantification of mammalian gene expression control", Nature 473(7347):337, DOI: 10.1038/nature10098 (link)
- Liu, Y. et al., (2016), "On the dependency of cellular protein levels on mRNA abundance", Cell, 165(3):535, DOI: 10.1016/j.cell.2016.03.014 (link)
- Jovanovic, M. et al., (2015), "Dynamic profiling of the protein life cycle in response to pathogens", Science, 347(6226):1259038, DOI: 10.1126/science.1259038 (link)
- Vogel C. et al., (2012), "Insights into the regulation of protein abundance from proteomic and transcriptomic analyses" Nat Rev Genet, 13(4):227, DOI: 10.1038/nrg3185 (link)
- Payne, SH. et al., (2015), "The utility of protein and mRNA corrleation", Trends Biotechnol., 33(1):5, DOI: 10.1016/j.tibtech.2014.10.010 (link)
- Rodero, SH. et al., (2016), "Typ eI interferon-mediated monogenic autoinflammation: The type I interferonopathies, a conceptual overview", J Exp Med., 213(12):2527, DOI: 10.1084/jem.20161596 (link)
- Hardy, MP., et al., (2004), "Characterization of the type I interferon locus and identification of novel genes", Genomics, 84(2):331, DOI: 10.1016/j.ygeno.2004.03.003 (link)
- Morand EF, et al., (2020), "Trial of Anifrolu-mab in Active Systemic Lupus Erythematosus", N Eng J Med 382(3):211, DOI: 10.1056/NEJMoa1912196 (link)
- Bastard P. et al., (2020), "Autoantibodies against type I IFNs in patients with life-threatening COVID-19", Science 370(6515):eabd4585 DOI: 10.1126/science.abd4585 (link)
- Bastard P. et al., (2021), "Autoantibodies neutralizing type I IFNs are present in ~4% of uninfected individuals over 70 years old and account for ~20% of COVID-19 deaths", Sci Immunol., 6(62):eabl4340, DOI: 10.1126/sciimmunol.abl4340 (link)