The intricate bidirectional communication between interferons (IFNs) and the human microbiome represents a critical regulatory network that modulates host immunity, microbial ecology, and pathophysiological processes. The human microbiome, comprising trillions of microbial cells, functions as a metabolically active organ that profoundly influences host physiology through complex molecular interactions. Simultaneously, the interferon system serves as a primary mediator of innate immunity, coordinating antiviral responses and maintaining tissue homeostasis through JAK-STAT signaling cascades.
Recent advances have revealed extensive crosstalk between these systems, with significant implications for understanding immune regulation, disease pathogenesis, and therapeutic intervention strategies. This article highlights current understanding of IFN-microbiome interactions across different anatomical sites and discusses emerging therapeutic paradigms targeting this axis.
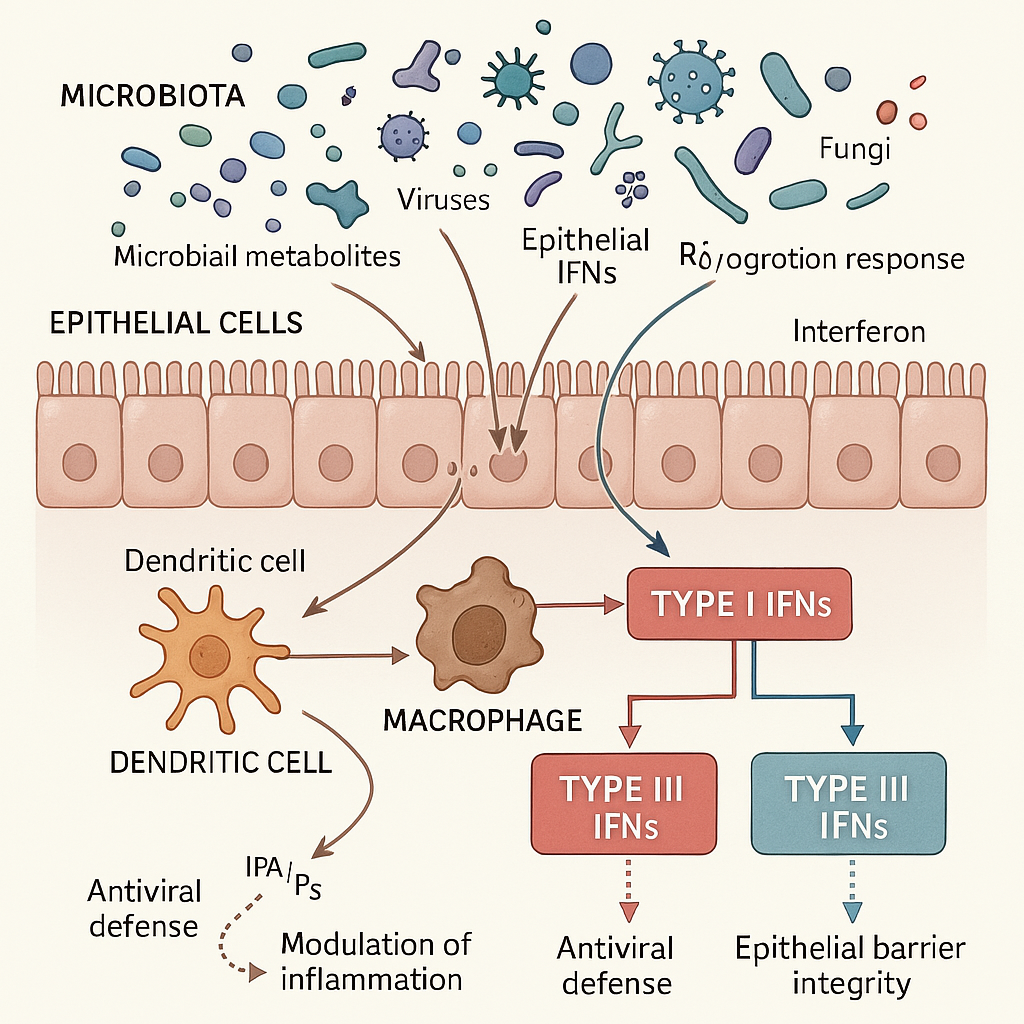
Type I Interferon-Microbiome Interactions
Molecular Mechanisms
Type I IFNs (e.g. IFN-α, IFN-β) exert pleiotropic effects on microbial communities through both direct antimicrobial activity and indirect modulation of the host immune microenvironment [1,2]. Specifically, González-Navajas et al. demonstrated that Type I IFNs regulate immune homeostasis through multiple mechanisms, including the enhancement of dendritic cell function, the promotion of T cell survival and differentiation, and the modulation of B cell responses, while also exhibiting context-dependent immunosuppressive effects through the induction of regulatory T cells and anti-inflammatory cytokines [1]. McNab et al. further elaborated that Type I IFNs have diverse effects on innate and adaptive immune cells during infection with viruses, bacteria, parasites, and fungi, directly and/or indirectly through the induction of other mediators, with their effects being highly dependent on the timing, duration, and cellular source of IFN production [2].
These interferons may contribute epithelial turnover and repair mechanisms, strengthening the physical barrier that separates commensal microorganisms from underlying immune effector cells. Saxena et al. revealed a paradoxical finding where human intestinal enteroids exposed to enteric viruses showed robust transcriptional interferon responses but limited functional antiviral activity, suggesting that the intestinal epithelium has evolved specialized mechanisms to balance antiviral defense with maintenance of barrier function and tolerance to commensal microbes [3].
The role of type I IFNs in bacterial infections is complex and context-dependent. Boxx and Cheng reviewed that type I IFNs can have variable effects on host susceptibility to bacterial infections, with outcomes that can be either protective or deleterious depending on the pathogen and host factors, including the bacterial species (intracellular vs. extracellular), infection route, timing of IFN response, and host genetic background [4]. In the gastrointestinal tract, type I interferons are implicated in both promoting innate immune defenses and modulating immune responses through anti-inflammatory cytokine production, though the extent and mechanisms of this dual role are not fully defined.
Recent evidence suggests commensal bacteria can enhance type I interferon responses, though the specific molecular pathways require further characterization. Zhang et al. demonstrated that commensal bacteria promote type I interferon signaling to maintain immune tolerance in mice [5]. Understanding these mechanisms remains an active area of investigation.
Pathophysiological Implications
Dysregulation of Type I IFN responses contributes to microbiome-associated pathology. Stefan et al. demonstrated that commensal microbiota could modulate natural resistance to virus infection, establishing complex feedback loops that influence both local immune responses and systemic immunity [6]. Imbalances in Type I IFN signaling have been associated with inflammatory conditions and shifts in microbial composition, yet the precise mechanisms connecting IFN dysregulation to microbiome-related pathologies are still under active investigation.
Type II Interferon-Mediated Microbiome Regulation
IFN-γ Signaling Pathways
IFN-γ represents the sole Type II interferon and functions as a master regulator of Th-1 mediated immunity. IFN-γ may influence microbiome through several proposed mechanisms, including macrophage activation, IDO-mediated tryptophan metabolism, and potential modulation of antimicrobial peptide expression, though direct evidence linking these pathways to microbiome alterations remains limited. Schoenborn and Wilson detailed that IFN-γ regulation occurs during both innate and adaptive immune responses through complex transcriptional and post-transcriptional mechanisms, including chromatin remodeling and microRNA-mediated control [7]. Macmickin reviewed that IFN-γ induces interferon-inducible effector mechanisms in cell-autonomous immunity, including autophagy, immunity-related GTPases, reactive nitrogen and oxygen intermediates, and nutritional immunity pathways that collectively create a hostile intracellular environment for pathogens [8].
IFN-γ plays a critical role in antimicrobial immunity by inducing a variety of physiologically significant responses. Boehm et al. demonstrated that treatment of cells with IFN-γ leads to changes in the expression of several hundred target genes that contribute to immune defense mechanisms, including MHC class I and II molecules, antigen processing machinery, and various antimicrobial effectors [9]. Tau and Rothman further elaborated that the cytokine exhibits immunomodulatory effects by enhancing antigen processing and presentation, increasing leukocyte trafficking, inducing an anti-viral state, and boosting anti-microbial functions through activation of the JAK1/JAK2-STAT1 signaling pathway [10].
Pathogen-Specific Responses
IFN-γ exhibits selective antimicrobial activity against intracellular pathogens, including Salmonella, Listeria, and Mycobacterium species. Neighbors et al. showed that early, innate production of IFN-γ is a critical step in immunological defense against certain intracellular bacterial pathogens, with IL-18 playing a crucial complementary role that extends beyond its effects on IFN-γ production [11]. This selectivity results from enhanced phagolysosomal fusion, increased nitric oxide production, and upregulation of immunity-related GTPases (IRGs) in infected macrophages.
Bao et al. demonstrated that in T cell-associated intestinal inflammation, IFN-γ plays an essential role in disease pathogenesis against Salmonella typhimurium infection, showing that IFN-γ-deficient mice exhibited increased bacterial colonization and systemic dissemination, demonstrating its complex functions in both protective immunity and inflammatory pathology [12]. The preservation of commensal populations during pathogen clearance suggests sophisticated targeting mechanisms, though the precise molecular basis for discrimination between beneficial and harmful microorganisms remains an area of active research.
Type III Interferon Functions at Mucosal Interfaces
Epithelial-Specific Signaling
Type III IFNs (IFN-λ1-4) demonstrate restricted cellular tropism, primarily targeting epithelial cells through the IFN-λR1/IL-10R2 receptor complex. Kotenko et al. first characterized that IFN-λs mediate antiviral protection through a distinct class of receptors composed of IFN-λR1 (IL-28Rα) and IL-10R2 subunits, which are primarily expressed on epithelial cells [13]. Sommereyns et al. further demonstrated that this specificity makes them ideally suited for mucosal immunity, where IFN-λ is expressed in a tissue-dependent fashion and primarily acts on epithelial cells in vivo, functioning as highly specialized cytokines uniquely suited to mediate antiviral protection at mucosal barriers [14].
Type III interferons play a prominent role in antiviral defense at respiratory and intestinal barrier surfaces, particularly through their action on epithelial cells. Lazear et al. comprehensively reviewed that they provide antiviral properties to cells by inducing the expression of hundreds of interferon-stimulated genes (ISGs), with many ISGs being commonly induced by both Type I and Type III IFNs, though with distinct kinetics and tissue distribution patterns [15]. However, recent evidence suggests that type III interferons can also disrupt epithelial barrier function under certain conditions, particularly during viral recognition and recovery from infection. Broggi et al. showed that IFN-λ suppresses intestinal inflammation by non-translational regulation of neutrophil function, demonstrating dual protective and potentially harmful effects [16]. Major et al. further revealed that Type I and III interferons can disrupt lung epithelial repair during recovery from viral infection, indicating that prolonged interferon responses may impair tissue healing [17].
Microbiome Modulation
Unlike Type I and II interferons, Type III IFNs demonstrate complex interactions with the microbiome. Stockmann et al. revealed through sophisticated spatial analysis that bacterial microbiota can stimulate homeostatic interferon-lambda responses within discrete pockets of intestinal epithelium, providing preemptive antiviral defense through recognition of bacterial products by epithelial pattern recognition receptors [18]. Conversely, Schott et al. demonstrated that certain microbiome compositions can impair interferon lambda production, as shown in oral mucosal tissues where specific bacterial communities suppress IFN-λ responses through mechanisms that remain incompletely characterized [19].
Baldridge et al. demonstrated that commensal microbes and interferon-λ work together to determine the persistence of enteric murine norovirus infection, showing that specific bacterial taxa can enhance IFN-λ responses [20]. Additionally, Hernández et al. showed that interferon-λ and interleukin 22 act synergistically for the induction of interferon-stimulated genes and control of rotavirus infection, indicating complex cytokine networks in gut immunity [21].
Pervolaraki et al. showed that the balance between type I and III interferons in the gut represents a tight regulatory mechanism between host protection and immunopathology, with different dependencies on mitogen-activated protein kinases to mount antiviral states in human gut tissues [22]. During enteric infections, the interplay between type I and III interferons contributes to antiviral immunity and may influence the maintenance of commensal populations, though the mechanisms underlying this balance are still being investigated.
Bidirectional Regulatory Networks
Microbiome-Derived Metabolites
The microbiome influences interferon responses through the production of bioactive metabolites, though this area requires further mechanistic investigation. Short-chain fatty acids (SCFAs) and other bacterial metabolites may modulate interferon signaling through various receptor-mediated pathways, but the specific molecular mechanisms and clinical relevance of these interactions need additional research to be fully characterized. These potential metabolite-mediated interactions could establish complex regulatory networks that influence both local and systemic immune responses.
Pattern Recognition Receptor Interactions
Commensal bacteria engage pattern recognition receptors (PRRs) including Toll-like receptors (TLRs) and nucleotide-binding oligomerization domain (NOD)-like receptors, potentially influencing baseline interferon tone. Ganal et al. demonstrated that commensal microbiota provides instructive signals that prime natural killer cells by nonmucosal mononuclear phagocytes, which may indirectly influence interferon responses and determine immunological outcomes during infections [23]. However, the direct effects of specific commensal bacterial products on PRR-mediated interferon production require further investigation.
Clinical and Therapeutic Implications
Current Understanding and Limitations
The interferon-microbiome axis offers potential opportunities for disease stratification and treatment monitoring, though clinical validation remains limited. The complexity of these interactions presents significant challenges for clinical translation. The variable effects of interferons on different bacterial species and the context-dependent nature of these responses complicate therapeutic targeting strategies. Individual variation in both interferon responses and microbiome composition suggests the need for personalized approaches, but standardized protocols and validated biomarkers are still under development.

Emerging Therapeutic Approaches
Several therapeutic approaches are being investigated that may target the interferon-microbiome axis:
Microbiome-Based Interventions: Rational design of interventions that could potentially enhance beneficial interferon responses while suppressing pathological inflammation remains largely theoretical. Targeted probiotic approaches and microbiome modulation strategies are areas of active research but lack robust clinical validation.
Combination Approaches: Integration of interferon-based therapies with microbiome-supporting interventions represents a promising but largely untested therapeutic strategy. Gopalakrishnan et al. demonstrated that gut microbiome composition modulates response to anti-PD-1 immunotherapy in melanoma patients through effects on antigen-presenting cells and T cell responses, suggesting that microbiota can influence interferon-dependent immune responses affecting treatment outcomes [24]. This provides proof-of-concept for microbiome-immune system interactions in clinical settings.
Precision Medicine Applications: Understanding how genetic polymorphisms in interferon signaling pathways interact with specific microbial signatures may help predict treatment outcomes, but such approaches remain largely investigational.
Future Directions and Challenges
Research Directions
Future research may focus on:
- Mechanistic characterization of cell-type-specific responses within complex tissue environments
- Development of sophisticated in vitro and in vivo models of interferon-microbiome interactions
- Integration of multi-omics approaches to understand system-level behavior
- Clinical validation of biomarkers and therapeutic targets
- Standardization of microbiome analysis protocols across studies
Clinical Translation Challenges
Moving from mechanistic understanding to clinical application requires addressing several significant challenges, including standardization of microbiome analysis protocols, development of stable therapeutic formulations, design of clinical trials that account for substantial inter-individual variation in both immune responses and microbial communities, and establishment of regulatory frameworks for microbiome-based interventions.
Conclusion
The interferon-microbiome axis represents a fundamental regulatory network that integrates microbial ecology with host immunity. While our understanding of these interactions at the molecular, cellular, and systems levels is advancing, much remains to be discovered about their clinical significance and therapeutic potential. The complexity of these interactions necessitates interdisciplinary collaboration between immunologists, microbiologists, bioinformaticians, and clinicians.
References
[1] González-Navajas JM, Lee J, David M, Raz E. Immunomodulatory functions of type I interferons. Nat Rev Immunol. 2012;12(2):125-135. doi:10.1038/nri3133. Available at: https://www.nature.com/articles/nri3133
[2] McNab F, Mayer-Barber K, Sher A, Wack A, O'Garra A. Type I interferons in infectious disease. Nat Rev Immunol. 2015;15(2):87-103. doi:10.1038/nri3787. Available at: https://www.nature.com/articles/nri3787
[3] Saxena K, Simon LM, Zeng XL, et al. A paradox of transcriptional and functional innate interferon responses of human intestinal enteroids to enteric virus infection. Proc Natl Acad Sci U S A. 2017;114(4):E570-E579. doi:10.1073/pnas.1615422114. Available at: https://www.pnas.org/doi/10.1073/pnas.1615422114
[4] Boxx GM, Cheng G. The roles of type I interferon in bacterial infection. Cell Host Microbe. 2016;19(6):760-769. doi:10.1016/j.chom.2016.05.016. Available at: https://www.cell.com/cell-host-microbe/fulltext/S1931-3128(16)30212-8
[5] Zhang B, Chassaing B, Shi Z, et al. Commensal bacteria promote type I interferon signaling to maintain immune tolerance in mice. J Exp Med. 2024;221(1):e20230063. doi:10.1084/jem.20230063. Available at: https://rupress.org/jem/article/221/1/e20230063/214644/Commensal-bacteria-promote-type-I-interferon
[6] Stefan KL, Kim MV, Iwasaki A, Kasper DL. Commensal microbiota modulation of natural resistance to virus infection. Cell. 2020;183(5):1312-1324.e10. doi:10.1016/j.cell.2020.10.047. Available at: https://www.sciencedirect.com/science/article/pii/S0092867420314549
[7] Schoenborn JR, Wilson CB. Regulation of interferon-gamma during innate and adaptive immune responses. Adv Immunol. 2007;96:41-101. doi:10.1016/S0065-2776(07)96002-2. Available at: https://www.sciencedirect.com/science/article/pii/S0065277607960022
[8] MacMicking JD. Interferon-inducible effector mechanisms in cell-autonomous immunity. Nat Rev Immunol. 2012;12(5):367-382. doi:10.1038/nri3210. Available at: https://www.nature.com/articles/nri3210
[9] Boehm U, Klamp T, Groot M, Howard JC. Cellular responses to interferon-gamma. Annu Rev Immunol. 1997;15:749-795. doi:10.1146/annurev.immunol.15.1.749. Available at: https://www.annualreviews.org/doi/10.1146/annurev.immunol.15.1.749
[10] Tau G, Rothman P. Biologic functions of the IFN-gamma receptors. Allergy. 1999;54(12):1233-1251. doi:10.1034/j.1398-9995.1999.00099.x. Available at: https://onlinelibrary.wiley.com/doi/10.1034/j.1398-9995.1999.00099.x
[11] Neighbors M, Xu X, Barrat FJ, et al. A critical role for interleukin 18 in primary and memory effector responses to Listeria monocytogenes that extends beyond its effects on Interferon-gamma production. J Exp Med. 2001;194(3):343-354. doi:10.1084/jem.194.3.343. Available at: https://rupress.org/jem/article-abstract/194/3/343/26036/A-Critical-Role-for-Interleukin-18-in-Primary-and
[12] Bao S, Beagley KW, France MP, et al. Interferon-gamma plays a critical role in intestinal immunity against Salmonella typhimurium infection. Immunology. 2000;99(3):464-472. doi:10.1046/j.1365-2567.2000.00955.x. Available at: https://onlinelibrary.wiley.com/doi/10.1046/j.1365-2567.2000.00955.x
[13] Kotenko SV, Gallagher G, Baurin VV, et al. IFN-lambdas mediate antiviral protection through a distinct class of receptors. Nat Immunol. 2003;4(1):69-77. doi:10.1038/ni875. Available at: https://www.nature.com/articles/ni87
[14] Sommereyns C, Paul S, Staeheli P, Michiels T. IFN-lambda (IFN-lambda) is expressed in a tissue-dependent fashion and primarily acts on epithelial cells in vivo. PLoS Pathog. 2008;4(3):e1000017. doi:10.1371/journal.ppat.1000017. Available at: https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.1000017
[15] Lazear HM, Schoggins JW, Diamond MS. Shared and distinct functions of type I and type III interferons. Immunity. 2019;50(4):907-923. doi:10.1016/j.immuni.2019.03.025. Available at: https://www.sciencedirect.com/science/article/pii/S1074761319301426
[16] Broggi A, Tan Y, Granucci F, Zanoni I. IFN-λ suppresses intestinal inflammation by non-translational regulation of neutrophil function. Nat Immunol. 2017;18(10):1084-1093. doi:10.1038/ni.3821. Available at: https://www.nature.com/articles/ni.3821
[17] Major J, Crotta S, Llorian M, et al. Type I and III interferons disrupt lung epithelial repair during recovery from viral infection. Science. 2020;369(6504):712-717. doi:10.1126/science.abc2061. Available at: https://www.science.org/doi/10.1126/science.abc2061
[18] Stockmann H, Kryczek I, Xiang L, et al. Homeostatic interferon-lambda response to bacterial microbiota stimulates preemptive antiviral defense within discrete pockets of intestinal epithelium. eLife. 2022;11:e74072. doi:10.7554/eLife.74072. Available at: https://elifesciences.org/articles/74072
[19] Schott C, Peterson LW, Rankin S, et al. Microbiome-mediated incapacitation of interferon lambda production in the oral mucosa. Proc Natl Acad Sci U S A. 2021;118(36):e2105170118. doi:10.1073/pnas.2105170118. Available at: https://www.pnas.org/doi/10.1073/pnas.2105170118
[20] Baldridge MT, Nice TJ, McCune BT, et al. Commensal microbes and interferon-λ determine persistence of enteric murine norovirus infection. Science. 2015;347(6219):266-269. doi:10.1126/science.1258025. Available at: https://www.science.org/doi/10.1126/science.1258025
[21] Hernández PP, Mahlakõiv T, Yang I, et al. Interferon-λ and interleukin 22 act synergistically for the induction of interferon-stimulated genes and control of rotavirus infection. Nat Immunol. 2015;16(7):698-707. doi:10.1038/ni.3180. Available at: https://www.nature.com/articles/ni.3180
[22] Pervolaraki K, Stanifer ML, Münchau S, et al. Type I and type III interferons display different dependency on mitogen-activated protein kinases to mount an antiviral state in the human gut. Front Immunol. 2017;8:459. doi:10.3389/fimmu.2017.00459. Available at: https://www.frontiersin.org/articles/10.3389/fimmu.2017.00459/full
[23] Ganal SC, Sanos SL, Kallfass C, et al. Priming of natural killer cells by nonmucosal mononuclear phagocytes requires instructive signals from commensal microbiota. Immunity. 2012;37(1):171-186. doi:10.1016/j.immuni.2012.05.020. Available at: https://www.cell.com/immunity/pdf/S1074-7613(12)00243-9.pdf
[24] Gopalakrishnan V, Spencer CN, Nezi L, et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science. 2018;359(6371):97-103. doi:10.1126/science.aan4236. Available at: https://www.science.org/doi/10.1126/science.aan4236